小麦的染色体数为42条。下图表示小麦的三个纯种品系的部分染色体及基因组成:Ⅰ、Ⅱ表示染色体,A为矮杆基因,B为抗矮黄病基因,E为抗条斑病基因,均为显性。乙品系和丙品系由普通小麦与近缘种偃麦草杂交后,经多代选育而来(图中黑色部分是来自偃麦草的染色体片段)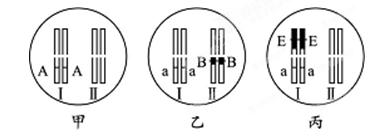
(1)乙、丙品系在培育过程中发生了染色体的 ____________变异。该现象如在自然条件下发生,可为生物进化提供原材料。
(2)甲和乙杂交所得到的F1自交,所有染色体正常联会,则基因A与a可随同源染色体的分开而分离。F1自交所得F2中有9种基因型,其中仅表现抗矮黄病的基因型有________种。
(3)甲与丙杂交所得到的F1自交,减数分裂中Ⅰ甲与Ⅰ丙因差异较大不能正常配对,而其它染色体正常配对,可观察到__________个四分体;该减数分裂 正常完成,可生产 ________种基因型的配子,配子中最多含有________条染色体。
正常完成,可生产 ________种基因型的配子,配子中最多含有________条染色体。
(4)让(2)中F1与(3)中F1杂交,若各种配子的形成机会和可育性相等,产生的种子均发育正常,则后代植株同时表现三种性状的几率为__________。
科学家在研究DNA分子复制方式时进行了如下的实验研究(已知培养用的细菌大约每20min分裂一次,产生子代,实验结果见相关图示):

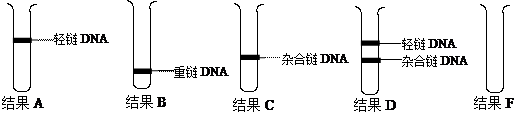
(1)实验一、实验二的作用是_______________________________________。
(2)从结果C、D看,DNA复制具有__________的特点。根据这一特点,理论上结果E中含14N的DNA分子所占比例为__________。
(3)复制过程除需要模板DNA、脱氧核苷酸外,还需要____________等条件。
(4)若对结果C中的DNA分子先用解旋酶处理,然后再离心,结果为F,请在图中表示出。
(5)若某次实验的结果中,结果C比以往实验结果所呈现的带略宽,可能的原因是新合成DNA单链中的N尚有少部分为____________。
(6分)某同学进行实验,甲图为实验开始状态,乙图为实验结束状态。请在乙图所示实验结果的基础上继续实验,探究蔗糖的水解产物能否通过半透膜。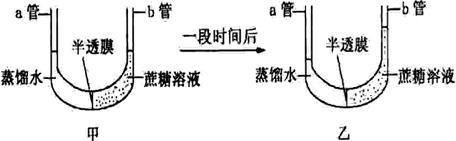
增添的实验材料:蔗糖酶溶液、斐林试剂、试管、滴管、水浴锅等。
⑴设计出继续实验的简要步骤:
① 向a管和b管分别加入等量的,水浴加热U形管至适宜温度,观察a和b两管内液面的高度变化;
② 吸取a和b两管内适量液体,分别加入A和B两支试管中,并加入斐林试剂,60~65℃水浴加热,观察A和B两支试管内有无。
⑵ 预测实验现象并作出结论。
① 如果a和b两管内液面的高度差 ,且A和B两支试管中
,则蔗糖的水解产物能通过半透膜;
② 如果a和b两管内液面的高度差 ,且A和B两支试管中
,则蔗糖的水解产物不能通过半透膜。
以下是两个实验,根据表格内容分析作答:
表1:探究温度对酶的活性(催化效率)的影响
| 操作步骤 |
操作方法 |
试管A |
试管B |
试管C |
| 1 |
加淀粉溶液 |
2mL[来 |
2 mL |
2 mL |
| 2 |
控制温度 |
60℃ |
100℃ |
0℃ |
| 3 |
加淀粉酶溶液 |
1 mL |
1 mL |
1 mL |
| 4 |
加碘液 |
2滴 |
2滴 |
2滴 |
⑴ 表1中的实验研究中的自变量是,因变量是。
⑵ 加碘液后如果溶液颜色越深,则说明酶的活性越 ________。
表2:探究酶催化作用的专一性
| 操作顺序 |
操作方法 |
试管A |
试管B |
| 1 |
加淀粉溶液 |
2 mL |
|
| 2 |
加蔗糖溶液 |
2 mL |
|
| 3 |
加斐林试剂甲 |
2 mL |
2 mL |
| 4 |
加斐林试剂乙 |
数滴 |
数滴 |
| 5 |
加淀粉酶溶液 |
1 mL |
1 mL |
⑶ 表2实验中存在三点错误,请用简要文字加以修正:
①______________ ___②____ _____
③_____
人的红细胞是两面凹的平扁盘状,红细胞中含血红蛋白,能携带氧气。血红蛋白由四条多肽链组成,其中2条α链各有141个氨基酸,另2条β链各有146个氨基酸。哺乳类包括人在内,成熟的红细胞都没有细胞核。但红细胞发育早期是有核的,以后细胞中血红蛋白分子迅速增加,达到红细胞干重的90%,细胞核以及线粒体、高尔基体、内质网和核糖体等从细胞中排除出去,红细胞变成一个富含血红蛋白的无核细胞而进入血液循环中去。
请回答下列问题:
⑴ 人类红细胞所需的能量主要来自于细胞内的哪类化合物?。葡萄糖由血浆进入红细胞的方式为。
⑵ 一个血红蛋白分子中含有个肽键。
⑶ 若把人的红细胞置于清水中,会发生的现象与结果是;若把人的红细胞置于浓盐水中,会发生的现象与结果又是。
⑷ 利用红细胞制备细胞膜进行成分分析,需要用(填方法)才能获得较为纯净的细胞膜。
请根据下列反应式回答:
⑴ 上述反应从左向右进行时,能量来自于ATP分子中的。
⑵ ATP分子中,A代表 ,由两种小分子组成。
⑶ 在各种生物的细胞中,ADP转化为ATP时所需的能量,可来自,也可来自。