下面是一个细胞的亚显微结构模式图,据图回答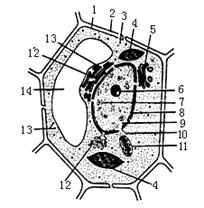
(1)该细胞是高等植物细胞,而不是低等植物细胞,判断的依据是 。
(2)蛋白质是在[ ] 合成的,为这一生命活动提供能量的细胞器是[ ] 。
(3)能够保证植物细胞坚挺的细胞器是[ ] 。
(4)含有色素的细胞器有[ ] 和[ ] 。
(5)结构10实现了核质之间频繁的 和 。
(6)结构1的主要成分是 。
(7)简要指出动物细胞与该细胞在结构上的差异 。
概念图是一种能够比较直观体现概念之间关系的图示方法,请根据所学知识,完成有关“生物体的结构”相关的概念图。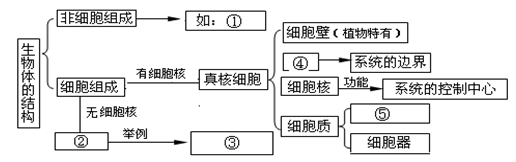
① ② ③ ④ ⑤
下图是细胞亚显微结构的部分模式图,请据图回答下列问题:(在[ ]内填写序号,后面横线填写名称)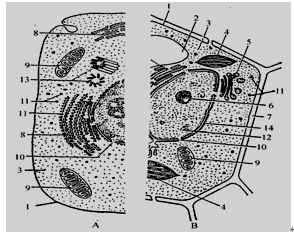
⑴图中的B半图是细胞,因为它具有[ ],[ ]和主要由纤维素构成的[ ]等结构。[1]所示结构的功能特点是具有。
⑵细胞生命活动所需能量主要来自[ ]。该结构主要进行的生理活动是。
⑶能将光能转化为化学能,并合成有机物的细胞器是[ ]。
⑷“生产蛋白质的机器”是 [ ],与细胞有丝分裂有关的细胞器是[ ]。
(7分)刚刚收获的甘薯与贮存一段时间的甘薯,甜度不同,贮存的甘薯明显变甜。有人推测其原因可能是贮存期间甘薯细胞内的非还原糖转变为还原糖。若用探究实验的方法证明这个推测,请回答下列有关问题。
(1)还原糖的鉴定,最常用的试剂是________,鉴定结果若出现____________现象,表明被测样品中含有还原糖。
(2)请结合提示,完成实验的设计过程。
①将刚收获的甘薯适量与已经贮存一段时间的等量甘薯,分别磨浆待用,标记为A、B浆液。
②取甲、乙两支试管分别加入等量的____________________________。
③在两支试管中同时加入_________________________。
④两支试管同时___________________________,观察现象。
⑤若___________________________,则证明推测成立。
⑥若_________________________________,则证明推测不成立。
根据下面图解,回答问题: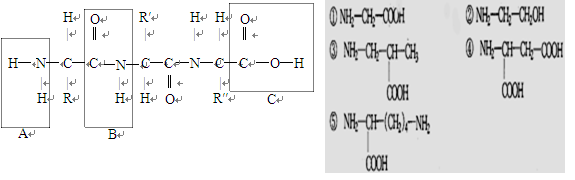
(1)左图中的化合物名称是,形成该化合物的生物化学反应叫做,它是由个氨基酸失去了分子水而形成的,含个肽键。
A是、B是、C是。氨基酸的结构通式可表示为
(2)右图5种化合物中属于氨基酸的是,这些氨基酸间的差别是不同。
右图是某生态系统中食物网简图,图中甲—庚代表各种不同的生物。请据图分析回答:
(1)此生态系统中作为生产者的生物是;作为次级消费者的生物是。
(2)生物甲与己的关系是。
(3)该生态系统只表示了部分成分,图中未表示的成分有、。
(4)生态系统中能量流动的特点是。