为研究根背光生长与生长素的关系,将水稻幼苗分别培养在含不同浓度生长素或适宜浓度NPA (生长素运输抑制剂)的溶液中,用水平单侧光照射根部(如下图),测得根的弯曲角度及生长速率如下表: 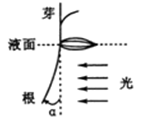
| 处理方式 测定指标 |
外源生长素(mg/L) |
NPA(μmol/L) |
|||
| 0 |
0.001 |
0.01 |
0.1 |
3 |
|
| 弯曲角度a(度) |
37 |
40 |
31 |
22 |
16 |
| 生长速率(mm/天) |
15 |
17 |
13 |
11 |
8 |
据此实验的结果,不能得出的结论是
A.根向光一侧的生长速率大于背光一侧
B.生长素对水稻根生长的作用具有两面性
C.单侧光对向光一侧生长素的合成没有影响
D.单侧光照射下根的背光生长与生长素的运输有关
细胞膜与其完成各种生理功能相适应,极为重要的结构特点
| A.磷脂排列成双分子层 | B.蛋白质分子覆盖或镶嵌于磷脂双分子层中 |
| C.两侧膜物质分子排列不对称 | D.膜物质分子的运动使其具有流动性 |
用紫色洋葱鳞片叶表皮细胞做质壁分离的实验,若发生质壁分离前观察的细胞如下图甲,则发生质壁分离时所观察到的细胞是乙图中的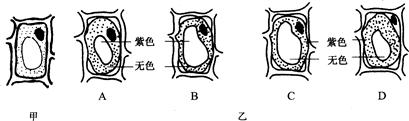
向鱼鳔(半透膜)内注入30%的蔗糖溶液,扎紧入口并称重,然后将其浸入5%的蔗糖溶液中,每隔半个小时称一次重量,结果如下图所示。下列关于其重量变化的解释正确的是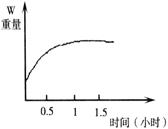
①在进行实验的1 小时之内,鱼鳔重量增加是因为外界水分进入鱼鳔内
小时之内,鱼鳔重量增加是因为外界水分进入鱼鳔内
②在进行实验的1小时之内,鱼鳔重量增加是因为外界蔗糖分子进入鱼鳔内
③在进行实验的1小时后,鱼鳔重量不再增加是因为进出鱼鳔的水分子处于相对平衡状态
④在进行实验的1小时后,鱼鳔重量不再增加,是因为进出鱼鳔的蔗糖分子处于相对平衡状态
| A.①② | B.②④ | C.①③ | D.②③ |
细胞作为最基本的生命系统,具有系统的一般特征:有边界,有系统内各组分的分工合作,有控制中心起调控作用。下列相关说法错误的是
| A.细胞的边界是细胞壁 |
| B.细胞内各种细胞器既有分工,又有合作 |
| C.功能不同的细胞在细胞器的种类、数量上存在差异 |
| D.系统的控制中心——细胞核,控制着细胞的代谢和遗传 |
细胞因某种原因改变了磷脂双分子层的排列,受到影响的细胞器或细胞结构是
| A.中心体 、高尔基体、 内质网、液泡 | B.液泡、叶绿体、线粒体、高尔基体 |
| C.细胞膜、叶绿体、线粒体、核糖体 | D.线粒体、核糖体、中心体、高尔基体 |