(6分) 下图甲示反射弧相关结构,图乙是图甲中某一结构的亚显微结构模式图。请分析回答: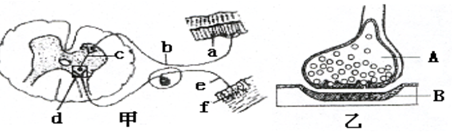
(1) 甲图中f表示的结构是_____ ____。
(2)乙图是甲图中___ ___(填字母)的亚显微结构放大模式图,称为 。
(3) 缩手反射时,兴奋从A传到B的信号物质是___________,传递的特点是 。
(4)甲图中该反射弧结构具有 个神经元。
日本下村修、美国沙尔菲和美籍华人钱永健因在研究绿色荧光蛋白(GFP)等方面的突出贡献,获得2008年度诺贝尔化学奖。GFP会发出绿色荧光,该蛋白质在生物工程中有着广阔的应用前景。请据图回答相关问题。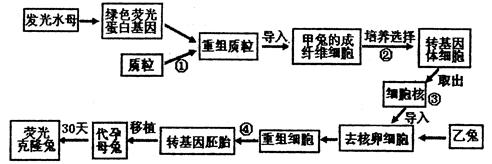
(1)绿色荧光蛋白基因在该实验中是做为基因。
(2)在②过程进行转基因体细胞的筛选时,比较简易的筛选方法是 。
。
(3)在④过程培养重组细胞时,培养 液中通常含有葡萄糖、氨基酸、无机盐、维生素等。
液中通常含有葡萄糖、氨基酸、无机盐、维生素等。
(4)以上培育过程中应用到的生物技术有等。
(5)在以上③核移植过程中,乙兔提供的细胞应是,移核之前要对此去核的原因是。
下图为哺乳动物的胚胎干细胞及其分化的示意图。请回答:
(1)胚胎干细胞是一类在功能上有 的细胞。
(2)图中分化程度最低的干细胞是______。在体外培养条件下,培养液中加入___因子,可诱导该种干细胞向不同类型的组织细胞分化。
(3)在机体内,皮肤干细胞分化成皮肤细胞是机体细胞中基因___________的结果。
(4)某患者不能产生正常的白细胞,通过骨髓移植可以达到治疗的目的,骨髓的治疗的实质是将上图的______________细胞移植到患者体内。
(5)若要克隆某种哺乳动物,从理论上分析,上述红细胞、白细胞神经细胞中不能选用作为供体的细胞是成熟的___________,其原因是________ ___。
科学家从某些能无限增殖的细胞的细胞质中分离 出无限增殖调控基因(prG),该基因能激发许多动物细胞的分裂,这为单克隆抗体的制备提供了更为广阔的前景。请回答以下相关问题:
出无限增殖调控基因(prG),该基因能激发许多动物细胞的分裂,这为单克隆抗体的制备提供了更为广阔的前景。请回答以下相关问题:
(1)利用细胞融合技术制备单克隆抗体,促进细胞融合可选择 方法。
(2)有人提出,可以直接通过基因工程导入该调控基因来制备单克隆抗体,其思路如右图
所示。请回答:
 |
①酶A是指 ,酶B是指 。
②对已导入重组质粒的II进行“检测”的目的是 ;
Ⅲ细胞具有的特点是 。
(3)也有人提出用细胞核移植技术构建重组细胞来生产单克隆抗体,试用相关的图示及文字表示该过程:
降钙素是一种多肽类激素,临床上用于治疗骨质疏松症等。人的降钙素活性很低,半衰期较短。某科学机构为了研发一种活性高、半衰期长的新型降钙素,从预期新型降钙素的功能出发,推测相应的脱氧核苷酸序列,并人工合成了两条72个 碱基的DNA单链,两条链通过18个碱基对形成部分双链DNA片段,再利用Klenow酶补平,获得双链DNA,过程如下图。
碱基的DNA单链,两条链通过18个碱基对形成部分双链DNA片段,再利用Klenow酶补平,获得双链DNA,过程如下图。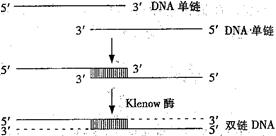
在此过程中发现,合成较长的核苷酸单链易产生缺失碱基的现象。分析回答下列问题:
(1)Klenow酶是一种________酶,合成的双链DNA有________个碱基对。
(2)获得的双链DNA经EcoRⅠ(识别序列和切割位点-G↓AATTC-)和BamHⅠ(识别序列和切割位点-G↓GATCC-)双酶切后插入到大肠杆菌质粒中,筛选含重组质粒的大肠杆菌并进行DNA测序验证。
①大肠杆菌是理想的受体细胞,这是因为它_________________________。
②设计EcoRⅠ和BamHⅠ双酶切的目的是___________________________。
③要进行重组质粒的鉴定和选择,需要大肠杆菌质粒中含有_______________。
(3)获得重组质粒如果导入到动物细胞中,最常用的方法是_______________。
(4)上述制备该新型降钙素,运用的现代生物工程技术是____________________。
下图表示用显微注射法导入目的基因获得转基因鼠的过程,据图回答。
(1)受精卵可以从供体母鼠体内获得,也可以通过体外受精获取。
①若受精卵细胞从供体母鼠体内获得,其基本过程是:首先对供体母鼠用促性腺激素进行_________处理,再通过自然交配或________完成受精,最后从供体母鼠输卵管中获得受精卵。
②若受精卵在体外通过体外受精获得,其基本操作过程包括________、________和受精。
③图中雌原核形成的同时在透明带和卵黄膜间会观察到 。
(2)过程Ⅲ表示受精卵经过体外培养,形成早期胚胎,再移植到受体母鼠体内。培养过程中,培养液添加一定量的 ,以防止杂菌污染;在含有5%CO2的培养箱中进行培养,CO2的主要作用是 。
(3)用显微注射技术将目的基因导入受体细胞,需要构建基因表 达载体,目的是____________
达载体,目的是____________
_ 。